




























Numerous factors or mechanisms may signal biological change; natural selection, genetic drift and other gene flow can all introduce variation into a population, but these are merely the effects, or end products of change, what physically lies at the source of the process - what environmental factor triggered or was driving the selection, drift and flow?
Responsiveness of Vertebrates to Environment
Many studies of vertebrates have suggested that climate is the single best indicator of modern species richness. Over the past decade several different theories have been suggested in hopes of establishing a clear and precise relationship between organismal change and the environment. Some of these hypotheses are relatively familiar, such as Van Valen’s “Red Queen model,” and Vrba’s “Turnover-pulse model,” but others less so… Regardless, the majority of these models work to resolve one key question; how synchronous are climate and biotic change?
Vertebrates, and in particular mammals, are highly responsive to changes in the environment. Climate fluctuation can lead to an incredible amount of variation in the diet choice, distribution and even morphology of vertebrates. Even within Humans, biological anthropologists have identified trends associated with body size and appendage length based on climate; they refer to this correlation as Bermand and Allan’s Rule. It suggests that human populations occupying regions closer to the equator statistically contain individuals with larger body size and longer limbs relative to those who inhabit more pole-ward regions. This variation has arisen primarily to promote more efficient thermoregulation as to reduce body heat loss in cooler regions, and to promote cooling in warmer climates. Some vertebrates posses even finer genetic plasticity changing morphologically with what could be termed “hair triggers.”
One of the most highly variable vertebrates is the Pocket gopher (Thomomys spp). These gophers have been intensively studied as indicators of environmental change. Their populations, and individual morphology, have been shown to vary with in a period of less than ten years. The most easily recognized change in morphology is their body size - which fluctuates with everything from altitude to diet. Body size, in turn, has a noticeable effect on territory, fertility of females and distribution of the population (Hadly, 1996). These physiological changes, and the fossils they have left behind, have been intensely studied in relation to Late-Holocene Climatic changes.

One site in particular, Lamar Cave, located with in the boundaries of Yellowstone National Park, has been used by Elizabeth Hadley to make correlations to such historic climate changes as the Medieval Warm Period (MWP). Through comparison of fossil dental plates with those of modern descendants, Hadley has been able to demonstrate that during the MWP pocket gophers of this region had the smallest body size than at any other point in the paleontological record. Using similar comparisons she was also able to identify trends in morphology that correlated with the Little Ice Age (LIA) and other events including neoglaciation (Hadly, 1996). The response of vertebrates is not limited to the highly plastic morphology of pocket gophers. Many other vertebrates respond equally as readily to climate change. However, morphological change is not necessarily required of a species in order for it to serve as a climate proxy. During their lifespan, all organisms make detailed records of their environment. Through the fundamental life processes of cellular growth and repair vertebrates have the potential to generate detailed records of their environment. Just as the old cliché exemplifies, “you are what you eat.” This record can later be resurrected and deciphered from their fossil remains to help clarify the picture of the climate and chemistry they endured while alive.
Isotopes
Examinations of isotopic records are commonplace for many paleontologists. Uncountable volumes have been written on Foram 18-O isotopes and their relationship to periods of increased glaciation, or similar correlations of speleothems and 13-C, but accurate isotopic records of both of these elements can be obtained from other sources as well - including vertebrates.
Carbonate apatite is a primary constituent of both skeletal bones and dentition in the vertebrates. It is incorporated into the organism through diet and water uptake. The process is riddled with various “vital effects” but has nonetheless been found to be very accurate. In fact, many conservationist use this isotopic signature in vertebrates to better decipher the life histories of contemporary vertebrates in hopes of aiding in their preservation. For example, remains from the world’s largest cavefish (Milyeringa veritas) have been used by Australian conservationists to better examine and comprehend the environment and ecosystem which the fish inhabits. The cavefish lives in deep columns of water that are rich in dissolved limestone. The depths of the water column posses various corresponding incremental concentrations of dissolved materials and by examining the isotopic signatures of the fish’s skeletal apatite, conservationists were able to discover the exact depth the fish occupies.
Milyeringa veritas

Uptake of elements within vertebrates (O, C, Sr & others) generates a highly detailed record of environmentally available isotopes, and as one might guess, availability is indicative of climate.
As another example, Thure Cerling and others at the University of Utah have refined the technique of laser ablation to such quality and precision that they can use it to vaporize the apatite found in vertebrate teeth, and then analyze the gas to determine its isotopic components. They can examine the teeth to such resolution as to be able to report the dietary preferences of mammals between C3 and C4 plants. Previously, samples of fossil teeth or skeletal structures were far to rare to permit them to be thrown into a TIMS for examination, but now that laser ablation has reached such a high level of efficiency isotopic analysis can be made in the smallest and rarest of specimens with minimum impact on the fossil itself. Laser ablation can be used to examine the minute layer of enamel on the smallest of rodents and the most voluble of fossils on display at museums with little difficulty.
As a side note - many scientists have recently discovered that skeletal tissues, which possess apatite, are very susceptible to diagenic processes that can cause recrystalization and alter results, but dental samples seem to be far more reliable when it comes to maintaining their original isotopic signatures. This coupled with the knowledge acquired in the past few decades in regards to the incorporation of specific 18-O and 13-C isotopes into biological entities has brought to light a highly detailed and accurate record of climate change.
Similar techniques have been used to examine bone fragments from marine vertebrates. Some alteration of samples occurs during fossil formation in paleo-seawater environments which forces users of this technique to integrate other proxies of isotopic variation in their final estimates as a “check,” but as the technique begins to advance some of this error is likely to be eliminated. Others have also verified the validity of the use of dental enamel as an isotopic record. Schmitz and his colleagues examined fossil shark teeth from two different locations in Mississippi (one strata being mixed marine, the other brackish-water). Both samples demonstrated nearly identical isotopic signatures, even though they had been deposited in different strata, one with a high salinity concentration - previously shown to accelerate diagenesis. They also examined skeletal remains from marine vertebrates, but there seemed to be a significant error associated with those found in the marine strata.
Isotopic signatures can be very useful in the interpretation of paleoclimate, but in certain cases the mere presence of an organism at a given location - biogeography - can report volumes of information in regards to past climate.
Biogeography
The vertebrate fossil record can be extrapolated in terms of the diversity, divergence and variation of distinct animal groups over a geographic region to display, with accuracy, the impact of environmental change. Similarly, known vertebrate biogeographic patterns can be used to demonstrate specific climatic fluctuation both locally and on the global scale.
The Cenozoic has been marked by tremendous variation of climate. During the early portions of the Cenozoic high temperatures were the standard with the highest to be found in the Early to Middle Eocene, since then a falling temperature trend has been the rule. It was during the Eocene-Oligocene transition that Antarctica became isolated and sea levels dropped dramatically with the accumulation of ice caps, this had the effect of changing terrestrial habitats significantly. Throughout the Cenozoic, similar patterns of fluctuating glacioeustatic sea level events would continue to occur. Evidence of this can be found across the globe.
Thailand, in South-East Asia, is one of many regions that accurately record these glacioeustatic events through its vertebrate fossil record. Thailand is especially significant in that it separates two distinct biogeographical regions (Fig 1). The Indochinese region is located to the north of Thailand and the Sundaic region is located to the south. These two regions have significantly different climates and zoological compositions. Up until about 800kyr B.P. fluctuation of sea level was somewhat constant in the region of Thailand, with the average being somewhere around 70m below the modern, and the greatest change being no more than 100m below the modern. However, near the 800kyr B.P. mark level dropped more dramatically, some fluctuations going as low as 170m below modern sea levels. This had the effect of constructing a land bridge between the Indochinese and Sundaic biogeographical regions, thus permitting the migration of a large variety of animals, including mammals. Through careful comparison of the fossils these migrating animals left behind, and the extant species found in both the Indochinese and Sundaic regions, certain conclusions can be established in regards to their phylogenies. The presence of fossil mammals with Indochinese affinity in the Sundaic regions, and the presence of Sundaic mammals in Indochina suggest some mode of migration in the past that is prevented in modern times by the sea acting as a barrier. Through radio dating of remains, an estimated time for the existence of the land bridge can be calculated, thus the fossil mammals provide dates for periods of reduced sea levels, and in turn, periods of increased glaciation.
FIGURE 1--- Land bridge linking distinct biogeographical regions in Thailand
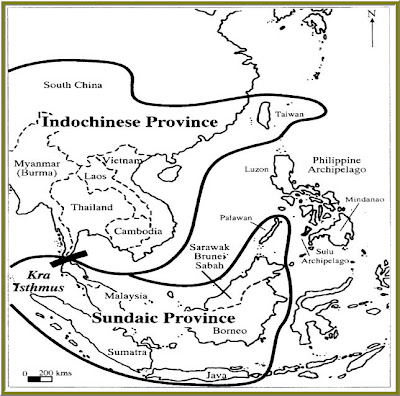 Thailand is not unique in its ability to provide vertebrate proxies of climate change. Similar scenarios have been played out across the globe. Grenada offers another example of vertebrate biogeography acting as an indicator of sea level change due to increased glaciation.
Thailand is not unique in its ability to provide vertebrate proxies of climate change. Similar scenarios have been played out across the globe. Grenada offers another example of vertebrate biogeography acting as an indicator of sea level change due to increased glaciation.Grenada has a rather sparse population of indigenous mammals at present, and bats represent the majority of these species. However, paleontologists have recently uncovered a variety of fossil mammals near the Grenadine coast dating from the Pliocene. These species, such as representative capybaras and sloth, did not arise independently on Grenada and then become extinct. Their lineages suggest that they arrived on Grenada from the South American mainland, where their descendants can still be found today. Grenada is located on the Southern Antilles Arc Platform, an elongate sub ocean structure that extends some 180km. This arc provides a base for several small islands and at its closest point comes to with in 40km of South America. During the Pliocene, a combination of low sea levels and tectonic activity provided a land bridge that may have conjoined Grenada with South America thus allowing for the migration of mammals. Through accurate dating of these fossil remains and of the associated tectonic activity estimates for late Pliocene glaciations can be made.
Through biogeographic research, other evidence can be in found with the target of determining periods of climatic change that are independent of sea level, and glacioeustaic events. Many other factors can have the effect of generating temporary corridors that permit faunal exchange. A “corridor” is simply a narrow tract, or pathway, which allows for the exchange of animals from one side to another. These corridors can be created by various means, including the growth of a forest.
The western Canadian ice-free corridor is a continuous tract of land that extends from Alberta, Northward to the Arctic Ocean (fig 2). This corridor separates Eastern and Central Canada from the Western Coastal portions of the Country. At one point in the past this corridor was closed to faunal exchange do to the advancing Laurentide ice sheet; however, near the close of the Pleistocene the ice retreated significantly enough to open the corridor temporarily, before finally closing again. This second closing of the corridor was not due to advancing ice caps, but rather to the growth of Boreal forests.
FIGURE 2---- Ice-free corridor, shaded region
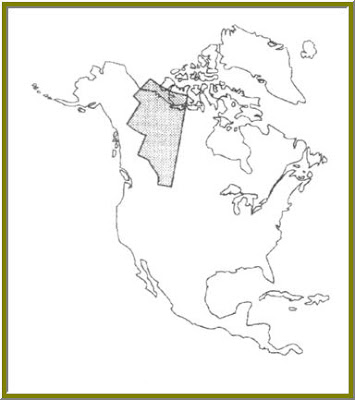
Interpretation of paleoclimate is a highly important field of study. Not only is it required to better understand the physical and biological history of our planet, but also to endow us with the knowledge needed to prepare for tomorrows’ inevitable changes. The Cenozoic has been marked by extreme change of temperature and climate as a whole, many such changes visible on a short-term scale. This period in geological history has seen everything from periods of hot and dry, to glaciation. To better understand the past and future effects on humans we must discover how it impacted the existence of other animals, in particular, other mammals. To better understand their evolutionary responses to climate, careful analysis of their fossil record must be made. Their physiology, morphology, biochemistry and even behavioral responses record every detail of their natural histories and the climates that they endured. Organisms change through time; the trick to rendering these records lays in the accurate translation of their fossil remains. To accomplish this end, the vertebrate indicators of climate change are an essential and indispensable tool and I very much look forward to reading more about them in May.
RECCOMENDED READING
Alberdi, M. et al. 2001. Vertebrate taphonomy in circum-lake environments: Three cases in the Guadix-Baza Basin (Granada, Spain). Paleogeogrphy, Paleoclimatology, Paleoecology. 165:1-26.
Badgley, C; Behrensmeyer, A. 1995. Preservational, Paleoecological and evolutionary patterns in the Paleogene of Wyoming-Montana and the Neogene of Pakistan. Paleogeography, Paleoclimatology, Paleoecology. 115: 319-340.
Badgley, C; Behrensmeyer, A. 1995. Two long geological records of continental ecosystems. Paleogeography, Paleoclimatology, Paleoecology. 115:1-11.
Barnosky, A. 2001. Distinguishing the effects of the Red Queen and the Court Jester on Pliocene Mammal Evolution in the Northern Rocky Mountains. Journal of Vertebrate Paleontolgy. 21:172-185.
Barrat, J; etal. 2000. Strontium isotopes in biogenic phosphates from a Neogene marine formation: implications for paleoseawater studies. Chemical Geology. 168:325-332.
Bromage, T.; etal. 1995. Paleobiography of the Molawi Rift: Age and vertebrate paleontology of the Chiwando beds, northern Molawi. Journal of Human Evolution. 28:37-57.
Bussuyt, F; Milankovitch, M. 2001. Amphibians as Indicators of Early Territory “ Out- of -India” Dispersal of Vertebrates. Science. 292:93-100.
Cerling, T; Sharp,Z. 1996. Stable carbon and oxygen isotope analysis of fossil tooth enamel using laser ablution. Paleogeography, Paleoclimatology, Paleoecology. 126:173-186.
Coope, G.R. et al. 1997. Climatic and Environmental Reconstructions based on fossil assemblages from Middle Devension (Weichselion) deposits of the river Thames at South Kensington, Central London, UK. Quaternary Science Reviews. 16:1163-1195.
Cruzan, M and Templeton, A. 2000. Paleoecology and Coalescence: phylographic analysis of hypothesis from the fossil record. TREE. 15:491-496.
Dennistan, R. etal. 1999. Integrating stalagmite, vertebrate and pollen sequences to investigate Holocene vegetation and climate change in the southern Midwestern United States. Quaternery Research. 52:381-387.
Flynn, J; Wyss, A. 1998. Recent advances in South American mammalian paleontology. TREE. 11:449-454.
Gao, C. et al. 2000. Last interglacial and Devension deposits of the river Great Ouse at Toolpack Form, Fenstanton, Cambrideshire, UK. Quarternary Science Reviews. 19:787-810.
Kerr, J; Packer, L. 1999. The environmental basis of North American species richness patterns among Epicauta (caleoptera: Meloidae). Biodiversity and Conservation. 8:617-628.
MacDonald, G; McLeod, T. 1996. The Holocene closing of the ‘ice-free’ corrider: A Biographical Perspective. Quarternary International. 32:57-95.
MacPhee, R.; Singer, R.; Diamond, M. 2000. Late Cenozoic land mammals from Grenada, lesser Antilles Island-Arc. American Museum Novitates. 3302:1-20.
Scasso, R.; Castro, L. 1999. Cenozoic phosphatic deposits in North Patagonia, Argentina: Phosphogenesis, sequence-stratigraphy and paleooceanography. Journal of South American Earth Sciences. 12:471-487.
Schmitz, B. et al. 1997. Testing 87Sr/86Sr as a paleosalinity indicator on mixed marine, brackish-water and terrestrial vertebrate skeletal apatite in late Paleocene-early Eocene near coastal sediments, Mississippi. Chemical Geology. 140:275-287.
Vigne, Jean-Denis. 1996. Small mammal fossil assemblages as indicators of environmental change in northern Corsica during the last 2500 years. Journal of Archeological Science. 23:199-215.
Wing, S; Alroy, J; Hickey, L. 1995. Plant and mammal diversity in the Paleocene to early Eocene of the Bighorn Basin. Paleogeography, Paleoclimatology, Paleoecology. 115:117-155.
Elizabeth A. Hadly (1997). Evolutionary and ecological response of pocket gophers (Thomomys talpoides) to late-Holocene climatic change Biological Journal of the Linnean Society, 60 (2), 277-296 DOI: 10.1111/j.1095-8312.1997.tb01496.x
William F. Humphreys (2001). Milyeringa veritus (Eleotridae), a remarkably versatile cave fish
from the arid tropics of Northwestern Australia.
Environmental Biology of Fishes, 62 (1/3), 297-313 DOI: 10.1023/A:1011880726946

No comments:
Post a Comment